Physical stimuli for vision are electromagnetic waves. The wavelength of the electromagnetic waves that a human can perceive as light ranges from about 400 to 700 nanometers (nm).
Humans have two types of receptors for vision. Rods and cones. Rods work under dim lighting conditions, whereas cones function under bright lighting conditions. Rods and cones are located on the retina, a thin layer in the back of the eye.
In the dark, we cannot see colors because there is only one kind of rods. We can distinguish objects only by the differences of brightness.
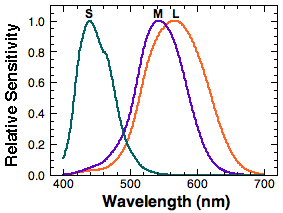
In the environment that is bright enough, we can see various colors. This is due to the three kinds of cones we have. They are called short-wavelength-sensitive (S) cones, middle-wavelength-sensitive (M) cones, and long-wavelength-sensitive (L) cones. Their spectral sensitivity, the sensitivity as a function of wavelength, varies depending on the type of cones. S cones are sensitive to the short wavelength region. M and L cones are sensitive to the middle to long wavelength regions. Note that the L cone and the M cone spectral sensitivities are close together compared to the rather isolated S cone sensitivity.
When the electromagnetic waves are absorbed by the photopigments in the cones, the cones generate neural signals. These signals are sent to various cells in the retina including ganglion cells. The retinal ganglion cells send their signals to the lateral geniculate nucleus (LGN) in the thalamus.
The first stage toward color perception is transduction of physical stimuli
into neural signals that takes place in the cones. In the second stage, the
neural signals from cones are combined to create one achromatic and two chromatic
mechanisms. The achromatic mechanism carries information on luminance (black
& white) and combines inputs from M and L cones. Two chromatic mechanisms
are L-M and S-(L+M) mechanisms. As the name implies, the L-M mechanism gets
inputs from L cones and negative inputs from M cones. The L-M mechanism is a
precursor to our perception of red and green. The S-(L+M) mechanism combines
positive inputs from S cones and negative inputs from the sum of L & M cone
signals. The signals from the S-(L+M) mechanism lead to our perception of blue
and yellow. These signals created in the second stage need to be further processed
in our cerebral cortex such as V1 and V4 in order for us to perceive colors
as we do.
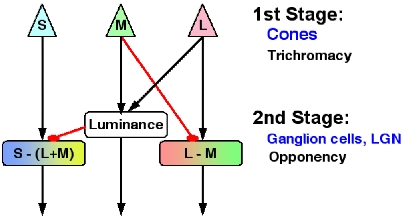
When we combine cone signals as described above, we can obtain output signals from the three mechanisms.
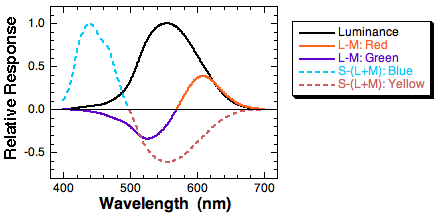
When we combine cone signals as described above, we can obtain output signals from the three mechanisms. The L-M function roughly corresponds to our perception of reds and greens. The S-(L+M) mechanism roughly corresponds to our perception of blues and yellows.
Majority of us have normal color vision. Some have defective color vision since birth and others develop color vision anomaly due to an injury to the brain or diseases. Congenital color vision defects have their origins in the genes and the condition is stable throughout one's life. People with normal color vision can lose color perception only, with other visual and cognitive functions intact, due to a specific lesion in the cerebral cortex. Such a condition is called cerebral achromatopsia. Other people may gradually develop reduced ability to perceive colors due to retinal diseases. We will focus on congenital color vision deficiencies here.
People with normal color vision are called normal trichromats because they possess three types of normal cone receptors (tri = three; chroma = color). There are other trichromats who also possess three types of cones, but one of the three types of cones has somewhat different spectral sensitivity compared to normal cones. They are collectively called anomalous trichromats. Protanomalous people have normal S and M cones and anomalous L cones whose spectral sensitivity is shifted to shorter wavelength (closer to the M cone spectral sensitivity). Deuteranomalous people have normal S and L cones and anomalous M cones whose spectral sensitivity is shifted to longer wavelength (closer to the L cone spectral sensitivity). Notice that the direction of the shift in the spectral sensitivity is always toward the other cone type. This attenuates the L-M signal, which leads to red-green color perception. Theoretically, there can be people whose S cone spectral sensitivity is shifted from that of normal S cones, but such a case is not confirmed solidly.
Trichromats have all the three types of functioning cones and should see all the different shades of colors. Normal trichromats are people with normal color vision. Anomalous trichromats have three types of cones, but one of the three types has a spectral sensitivity shifted from the normal cones. They see all the possible colors normal trichromats would see, but slightly differently. There are some people who entirely lack one type of cones. They are called dichromats and possess only two types of cones, instead of three. The dichromats who lack L cones are called protanopes, those who lack M cones are called deuteranopes, and those who lack S cones are named tritanopes. Because they lack one class of cones in their retinae, the shades of colors that they perceive are limited compared to trichromats.
There are small number of people who have only S cones or no cones at all. They are called monochromats. Because they do not have signals from different cone types to compare, they totally lack sensation of color.
The table below summarizes types of congenital color vision deficiencies. In the right-most column, the cone type with prime such as L' indicates that the person possess the type of cones whose spectral sensitivity is shifted from that of normal.
| Type | Condition | Person | Symbol | Cones |
|---|---|---|---|---|
| Anomalous | Protanomaly | Protanomal | PA | S M L' |
| trichromat | Deuteranomaly | Deuteranomal | DA | S M' L |
| Tritanomaly(?) | Tritanomal | TA | S' M L | |
| Dichromat | Protanopia | Protanope | P | S M |
| Deuteranopia | Deuteranope | D | S L | |
| Tritanopia | Tritanope | T | M L | |
| Monochromat | S-cone monochromatism | S-cone monochromat | S | |
| Rod monochromatism | Rod monochromat | None |
Among the people with congenital color vision defects, protans (protanomalous and protanopic individuals) have anomaly in L cones and deutans (deuteranomalous and deuteranopic individuals) have anomaly in M cones. Considering that the L-M mechanism leads to our perception of reds and greens and anything in between, we can easily infer that those people experience some difficulty in distinguishing theses colors, or lights in the middle to the long wavelength region. These defects are collectively called congenital red-green color vision deficiencies and are most prevalent forms of color vision defects. For example, the incidence of red-green color vision defects among caucasian males is about 8%.
The figure below is a comparison of the spectral sensitivities of L and M cones from normal trichromats (N), protanomalous trichromats (PA), and deuteranomalous trichromats (DA). Cone spectral sensitivities were taken from DeMarco, Pokorny, & Smith (1992).
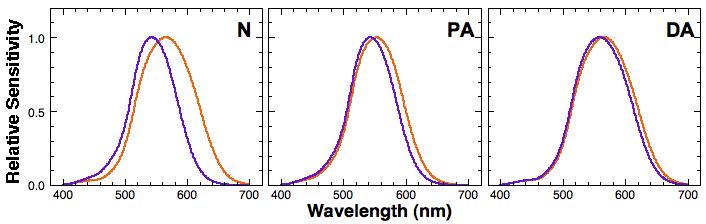
Notice the separation between the L and M cone spectral sensitivities among normal trichromatic, protanomalous, and deuteranomalous observers. The separation between the two in normal trichromats is about 30nm, whereas it is much smaller in protanomalous and deuteranomalous observers. This smaller separation translate into a smaller L-M signal when we calculate the difference between L and M cone signal for each wavelength. The figure below shows relative L-M signal (response) as a function of wavelength. Note that the L-M output by protanomalous and deuteranomalous observers is significantly attenuated compared to that of normal trichromats.
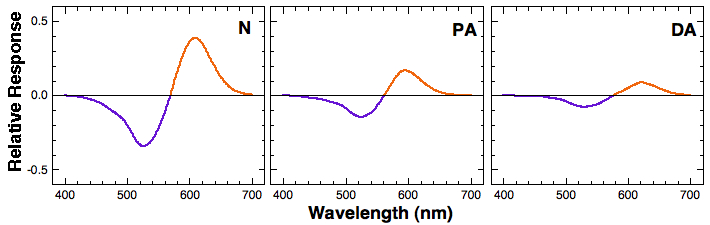
The attenuated L-M signal leads to a reduced sensitivity in discriminating colors in the middle- to long- wavelength region, which normally corresponds to colors green, yellow, orange, red, and brown. Protanomalous and deuteranomalous people would these colors slightly differently from normal trichromats. Or they may have some difficulty in distinguishing these colors. On the other hand, protanopes and deuteranopes entirely lack either L or M cones. This means that their visual system cannot form any L-M signal. Consequently, they cannot distinguish colors in the middle- to long- wavelength region at all. Practically speaking, however, some dichromats have learned to utilize other available cues such as luminance and the context to correctly "guess" colors.
Cones' spectral sensitivity is determined by the photopigments in them. The genes for the photopigments have been identified and mapped on the human chromosomes. L-cone-pigment genes and M-cone-pigment genes are located on the x-chromosome in tandem array.
DeMarco, P., Pokorny, J., & Smith, V.C. (1992). Full-spectrum cone sensitivity functions for X-chromosome-linked anomalous trichromats. Journal of the Optical Society of America A, 9, 1465-1476.